
Частная физиология ЦНС. Двигательные системы
Двигательные системы спинного мозга
Структурно-функциональная характеристика спинного мозга включает структуры, обеспечивающие формирование фазных и тонических (позных) рефлексов спинального организма.
Спинной мозг имеет сегментарное строение (31–33 сегмента) — каждый его сегмент связан с определенной частью тела. Спинной мозг включает пять отделов: шейный (СI–СVIII), грудной (ТhI–ТhXII), поясничный (LI–LV), крестцовый (SI–SV) и копчиковый (СоI–СоII).
В спинном мозге насчитывают около 13 млн нейронов (3% из них — мотонейроны, 97% — вставочные нейроны, относящиеся в том числе и к вегетативной нервной системе). При этом выделяют:
- нейроны соматической и вегетативной нервной системы;
- эфферентные, афферентные, вставочные, ассоциативные;
- возбуждающие и тормозные.
Центры спинного мозга участвуют в регуляции большинства внутренних органов и скелетных мышц, в частности центр зрачкового рефлекса локализуется в сегментах СVIII–ТhII, регуляции деятельности сердца — в ТhI–ТhV, слюноотделения — в ТhII–ТhV, регуляции функции желудка — в ТhVI–ТhIX, почек — в ТhV–LIII. Сегментарно расположены центры гладких мышц внутренних органов, центры пиломоторных рефлексов, центры, регулирующие функции потовых желез и сосудов. Парасимпатическую иннервацию получают из спинного мозга (SII–SIV) все органы малого таза: мочевой пузырь, часть толстой кишки ниже ее левого изгиба, половые органы. У женщин парасимпатическая иннервация обеспечивает сосудистые реакции клитора, влагалища; у мужчин — рефлекторный компонент эрекции.
Центры управления скелетной мускулатурой находятся во всех отделах спинного мозга и иннервируют по сегментарному принципу мышцы шеи (СI–СIV), диафрагмы (СIII–СV), верхних конечностей (СV–ТhII), туловища (ТhIII–LI) и нижних конечностей (LII–SV). Повреждения определенных сегментов спинного мозга или его проводящих путей вызывают специфические расстройства чувствительности и двигательные нарушения. Каждый сегмент спинного мозга участвует в чувствительной иннервации трех дерматомов. Имеется дублирование двигательной иннервации скелетных мышц, что повышает надежность функционирования опорно-двигательного аппарата и восприятия внешних раздражителей.
Ретикулярная формация спинного мозга обнаруживается на уровне шейных сегментов. Она участвует в регуляции тонуса мышц, вегетативных функций, афферентной им- пульсации.
Спинной мозг имеет собственный ассоциативный аппарат, устанавливающий связь между сегментами и внутри сегментов. Ассоциативный аппарат спинного мозга участвует в регуляции движений конечностей и туловища, тонуса мышц, поддержании позы.
Спинной мозг выполняет проводниковую и рефлекторную функции. Проводниковая функция спинного мозга и движения организма. С помощью проводниковой функции спинного мозга вышележащие отделы ЦНС получают информацию от организма и внутренних органов, управляют скелетной мускулатурой тела и конечностей, регулируют функции внутренних орга- нов.Проводниковая функция спинного мозга (табл. 4.1) осуществляется с помощью восходящих и нисходящих путей.Афферентная информация поступает в спинной мозг через задние корешки, эфферентная импульсация и регуляция функций различных органов и тканей организма осуществляются через передние корешки (закон Белла–Мажанди).
Таблица 4.1
Основные проводящие пути спинного мозга, обеспечивающие связь с вышележащими отделами ЦНС, и их роль
| Восходящие (чувствительные) пути | Физиологическое значение |
| Клиновидный пучок (пучок Бурдаха), проходит в задних столбах, импульса- ция поступает в кору | Проприорецептивная импульсация от верхней части туловища и рук |
| Тонкий пучок (пучок Голля), проходит в задних столбах, импульсация поступает в кору | Проприорецептивная импульсация от нижней части туловища и ног |
| Передний спинно-мозжечковый (Говерса) | Проприорецептивная импульсация поступает в мозжечок |
| Дорсальный спинно-мозжечковый (Флексига) | То же |
| Латеральный спинно-таламический | Болевая и температурная чувствительность |
| Передний спинно-таламический | Тактильная чувствительность — прикосновение, давление |
| Нисходящие (двигательные) пути | Физиологическое значение |
| Латеральный кортико-спинальный (пирамидный) | Импульсы к скелетным мышцам, произвольные движения |
| Передний кортико-спинальный (пирамидный) | То же |
| Рубро-спинальный (Монакова), проходит в боковых столбах | Участие в регуляции тонуса скелетных мышц |
| Ретикуло-спинальный, передние столбы | Регуляция тонуса скелетных мышц с помощью возбуждающих и тормозящих влияний на α- и γ-мотонейроны, а также регулирующие состояние спинальных вегетативных центров |
| Вестибуло-спинальный, передние столбы | Регуляция тонуса мышц, поддержание позы и восстановление нарушенной позы |
| Текто-спинальный, проходит в передних столбах | Импульсы, обеспечивающие осуществление зрительных и слуховых двигательных рефлексов (рефлексов четверохолмия) |
Значение афферентной импульсации, поступающей в спинной мозг:
- несет информацию об изменениях окружающей среды;
- участвует в координационной деятельности ЦНС по управлению скелетной мускулатурой — при выключении афферентной импульсации от рабочего органа управление им становится несовершенным;
- способствует поддержанию тонуса ЦНС — при выключении афферентной импульсации уменьшается суммарная тоническая активность ЦНС;
- участвует в процессах регуляции функций внутренних органов.
Двигательные системы спинного мозга обеспечивают формирование тонических,фазных рефлексов и ритмических рефлексов (классификация рефлексов по характеру ответной реакции) — это сегментарные рефлексы. Надсегментарные рефлексы осуществляются только с помощью шейного отдела.
По рецепторам, раздражение которых вызывает рефлексы, выделяют проприоцептивные, висцероцептивные и кожные рефлексы (защитные). Рефлексы, возникающие с проприорецепторов, участвуют в формировании акта ходьбы и регуляции мышечного тонуса. Висцероцептивные рефлексы возникают с интерорецепторов (рецепторов внутренних органов) и проявляются в сокращениях мышц передней брюшной стенки, грудной клетки и разгибателей спины. Эти рефлексы изучают в эксперименте у спинального организма с перерезанным спинным мозгом ниже сегментов С3–С5, где локализуются центры диафрагмальных нервов. В этом случае сохраняется диафрагмальное дыхание (центры диафрагмальных нервов связаны с головным мозгом).
Фазные рефлексы конечностей спинального организма. Рефлексы конечностей (рис. 4.13) в клинической практике исследуют наиболее часто.
Фазные рефлексы — это однократное сгибание или разгибание конечности при однократном раздражении кожных рецепторов или проприорецепторов.
Сгибательные рефлексы конечностей. Рефлексы, возникающие с рецепторов кожи, полисинаптические, они имеют защитное значение. Сгибательные (как и разгибательные) рефлексы, возникающие с проприорецепторов, — моносинаптические.
Фазные проприоцептивные рефлексы участвуют в формировании акта ходьбы. По степени выраженности фазных сгибательных и разгибательных рефлексов определяют состояние возбудимости ЦНС и возможные ее нарушения. Поэтому в клинической практике исследуют несколько сгибательных фазных рефлексов — ахиллов (пяточный) рефлекс, подошвенный рефлекс — кожный рефлекс, локтевой рефлекс — проприоцептивный.
При возникновении фазных сгибательных рефлексов одновременно с возбуждением мотонейронов мышц- сгибателей происходит торможение мотонейронов мышц- разгибателей этой же конечности — иначе сгибание будет невозможным.
Разгибательные рефлексы конечностей — моносинап- тические, возникают только с проприорецепторов мышц- разгибателей.
Фазные разгибательные рефлексы, в отличие от фазных сгибательных рефлексов, возникают в ответ на однократное раздражение только мышечных рецепторов, например при ударе по сухожилию четырехглавой мышцы ниже надколенника. При этом наблюдается коленный разгибательный рефлекс вследствие сокращения четырехглавой мышцы (мотонейроны мышц-сгибателей во время разгибательного рефлекса тормозятся с помощью постсинаптического реципрокного торможения посредством вставочных тормозных клеток Реншоу). Рефлекторная дуга коленного разгибательного рефлекса замыкается в поясничных сегментах (LII–LIV). Фазные разгибательные рефлексы, как и сгибательные, участвуют в формировании акта ходьбы (перемещение тела в пространстве).
Ритмические рефлексы. Характерным примером ритмических рефлексов является шагательный рефлекс.
Шагательные рефлексы вызываются у спинальной собаки (перерезан спинной мозг ниже сегментов С3–С5 — центр диафрагмального нерва, что обеспечивает диафрагмальное дыхание и жизнь организму). Они вызываются однократным раздражением кожи задней конечности, что вызывает ее сгибание и разгибание противоположной конечности — перекрестный и разгибательный рефлекс. Затем согнутая конечность разгибается и опускается, а разогнутая — сгибается и поднимается и т.д. Конечности не касаются опоры (собака зафиксирована в станке с помощью лямок), и без дополнительного раздражения конечности продолжают ритмично поочередно сгибаться и разгибаться — «шагать» в воздухе. Шагательный рефлекс (локомоция) может продолжаться часами, т.к. устранено тормозное влияние коры большого мозга. Импульсы к центрам сгибания и разгибания конечностей поступают от их проприорецепторов.
Роль рецепторов
Мышечные веретена (мышечные рецепторы, они первичные) расположены параллельно скелетной мышце — своими концами они крепятся к соединительнотканной оболочке (перимизию) пучка экстрафузальных мышечных волокон с помощью напоминающих сухожилия полосок соединительной ткани. В связи с этим, когда мышца расслабляется (удлиняется), растягиваются (раздражаются) и мышечные рецепторы, что и ведет к их возбуждению.
Сухожильные рецепторы (рецепторы Гольджи) локализуются в сухожилиях скелетных мышц, т.е. крепятся относительно скелетной мышцы последовательно, что обеспечивает их раздражение при сокращении мышцы и натяжении сухожилия. Поэтому сухожильные рецепторы посылают в мозг информацию о том, что мышца сокращена (напряжено и сухожилие), а мышечные рецепторы — что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра-антагониста (у мышц-сгибателей это возбуждение выражено слабее).
Механизм шагательного рефлекса. Когда мышца (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мышечные рецепторы. Импульсы от них поступают к своим α-мотонейронам спинного мозга и возбуждают их (рис. 4.14).
Поэтому α-мотонейроны, в свою очередь, посылают импульсы к этой же скелетной мышце, что ведет к ее сокращению. В результате сокращения мышцы возбуждение мышечных веретен прекращается или сильно ослабляется (они уже не растянуты), начинают возбуждаться сухожильные рецепторы, поскольку сухожилие натягивается (рис. 4.14, Б). Импульсы от сухожильных рецепторов поступают тоже в свой центр в спинном мозге, но к тормозным клеткам. Возбуждение этих клеток вызывает торможение α-мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется.
Ее расслабление (удлинение) ведет снова к возбуждению мышечных рецепторов и α-мотонейронов — мышца вновь сокращается. Вследствие сокращения снова возбуждаются сухожильные рецепторы и тормозные клетки в спинном мозге, что ведет к очередному расслаблению скелетной мышцы и т.д.
Мышца поочередно сокращается и расслабляется в результате поступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процессы в равной степени относятся и к мышце-сгибателю, и к мышце-разгибателю, при этом расслаблениескелетной мышцы запускает механизмы ее сокращения, а сокращение скелетной мышцы активирует механизмы, расслабляющие мышцу, что наблюдается при ходьбе и беге или любом повторяющемся действии. При ходьбе и беге, если на одной ноге сокращены сгибатели, на другой ноге сокращаются разгибатели, иначе нарушится поза. Это обеспечивается поступлением афферентных импульсов от мышечных и сухожильных рецепторов и поочередным возбуждением и торможением центров-сгибателей и центров-разгибателей другой конечности — перекрестный разгибательный рефлекс.
Шагательные координированные движения возможны при отсутствии обратной афферентации от проприоре- цепторов. Их программа формируется в онтогенезе. Они осуществляются с помощью межсегментарных связей на уровне спинного мозга и генератора локомоции каждой конечности. О наличии межсегментарных связей на уровне спинного мозга свидетельствует также факт вовлечения в шагательный рефлекс всех четырех конечностей при достаточно длительном и сильном раздражении одной конечности. Произвольные движения конечностей после разрыва спинного мозга невозможны — нет связи с корой большого мозга.
Спотыкательный рефлекс. У спинального организма во время шагательного рефлекса можно наблюдать споты- кательный рефлекс — при движении ноги вперед и столкновении ее с препятствием она поднимается и переступает через препятствие.
Тонические рефлексы спинального организма (рефлексы позы). Сегментарные рефлексы позы — это тонические сгибательные и разгибательные рефлексы конечностей, обеспечивающие сохранение естественной позы. Они возникают при длительном растяжении мышц и возбуждении мышечных рецепторов. Тоническое сокращение скелетных мышц является фоновым для всех двигательных актов, осуществляемых с помощью фазных сокращений мышц. Изменение тонуса мышц конечностей свидетельствует о нарушениях в ЦНС. В положении стоя тоническое сокращение мышц-разгибателей предотвращает сгибание нижних конечностей и обеспечивает сохранение вертикальной естественной позы. Тоническое сокращение мышц спины удерживает туловище в вертикальном положении, обеспечивая осанку человека. Тонические рефлексы на растяжение мышц (сгибания и разгибания) называют также миотатическими.
Таким образом, главная роль тонических рефлексов — поддержание естественной позы организма.
Надсегментарные рефлексы позы спинального организма — это шейные рефлексы. Они заключаются в перераспределении тонуса мышц конечностей при изменении положения шеи относительно туловища. Эти рефлексы осуществляются с участием различных отделов ЦНС. На уровне спинного мозга замыкаются шейные позные рефлексы, наличие которых установил голландский физиолог Р. Магнус (1924) в опытах на кошке. Имеется две группы этих рефлексов.
Первая группа шейных позных рефлексов возникает при наклоне головы вперед или назад. При наклоне головы вниз (кпереди) увеличивается тонус мышц-сгибателей передних конечностей и тонус мышц-разгибателей задних конечностей, в результате чего передние конечности сгибаются, а задние — разгибаются (рис. 4.15).
При наклоне головы вверх (кзади) возникают противоположные реакции — передние конечности разгибаются вследствие увеличения тонуса их мышц-разгибателей, а задние конечности сгибаются вследствие повышения тонуса их мышц-сгибателей. Эти рефлексы возникают с проприорецепторов мышц шеи и фасций,покрывающих шейный отдел позвоночника. В условиях естественного поведения животных они увеличивают шансы достать пищу, находящуюся выше или ниже головы, — это автоматизированная приспособительная реакция, рефлексы являются надсегментарными. Шейные рефлексы позы верхних конечностей у человека, по-видимому, утрачены, а рефлексы нижних конечностей выражаются не в сгибании или разгибании, а в перераспределении мышечного тонуса, обеспечивающего сохранение естественной позы.
Вторая группа шейных позных рефлексов (тонических) возникает с шейных проприорецепторов при поворотах или наклонах головы вправо или влево. Рефлекс выражается в повышении тонуса мышц-разгибателей обеих конечностей на стороне, в которую повернута (наклонена) голова, и в повышении тонуса мышц-сгибателей на противоположной стороне. Рефлекс направлен на сохранение позы, кото- раяможетбыть нарушенавследствиеизменения положения центра тяжести после поворота (наклона) головы. Центр тяжести смещается в сторону поворота головы — именно на этой стороне повышается тонус мышц-разгибателей обеих конечностей, что повышает устойчивость организма. Подобная ситуация наблюдается и у человека.
Спинальный шок. В случае повреждения спинного мозга у человека вследствие травмы наблюдаются мышечная атония и отсутствие рефлексов (спинальный шок). Главной причиной спинального шока является выключение влияния вышележащих отделов ЦНС на спинной мозг. Об этом свидетельствует тот факт, что повторная перерезка спинного мозга в эксперименте на животных ниже первой шока не вызывает. Спинальный шок у лягушек длится несколько минут, у собак — несколько дней, у человека — около 2 мес.
Тонус мышц спинального организма. После исчезновения спинального шока тонус мышц (иннервируемых посредством сегментов спинного мозга, которые находятся ниже перерезки) резко повышается. Сгибательные и разгибательные рефлексы нижних конечностей у человека в случае повреждения спинного мозга усиливаются (рис. 4.16).
Гипертонус имеет рефлекторную природу: деафферен- тация, например, нижней конечности у лягушки ведет к исчезновению тонуса у нее (опыт Бронжеста). Произвольные движения невозможны. Афферентная импульсация от мышечных рецепторов по задним корешкам спинного мозга (рис. 4.16), которая ведет к возбуждению α-мотонейронов спинного мозга, сокращению скелетных мышц и развитию гипертонуса, поддерживается двумя способами:
- спонтанной активностью мышечных рецепторов, что обеспечивает поступление импульсов от них к α-мотонейронам, возбуждение последних и сокращение мышц;
- спонтанной активностью γ-мотонейронов.
Возбуждение γ-мотонейронов вызывает возбуждение и сокращение иннервируемых ими интрафузальных мышечных волокон, в результате чего увеличивается натяжение мышечного рецептора, посколькуконцыегозафиксированы на скелетной мышце. Это вызывает раздражение и возбуждение мышечных рецепторов, импульсы от которых поступают к α-мотонейронам и возбуждают их. В свою очередь, α-мотонейроны посылают импульсы к скелетной мышце и вызывают ее постоянное (тоническое) сокращение, вследствие чего и развивается гипертонус у спинального организма. Если спинальное животное поставить на ноги, то оно будет стоять; если же положить на бок, оно будет лежать, не делая попытки встать на ноги. Функции центров спинного мозга управляются центрами головного мозга, и в частности ствола мозга. Животное с сохраненным средним мозгом называют мезэнцефальным организмом.
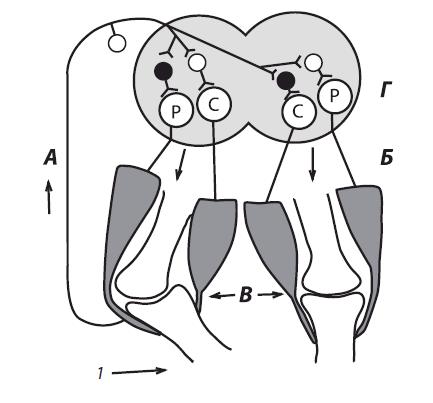
Рис. 4.13. Рефлексы нижних конечностей: А — дуга сгибательного (защитного) рефлекса; Б — дуга перекрестного разгибательного рефлекса; В — коленные суставы с мышцами; Г — сегмент спинного мозга; → — раздражение кожи; ↑ — афферентный путь; ↓ — эфферентные пути от α-мотонейронов центров сгибания (С) и разгибания (Р). Нейроны: светлые — возбуждающие; темные — тормозные
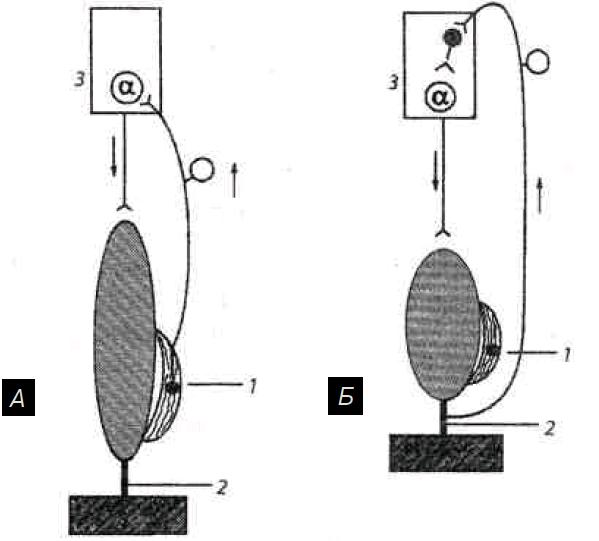
Рис. 4.14. Механизм возбуждения и торможения α-мотонейронов спинного мозга во время сокращения и расслабления скелетной мышцы: А — мышца растянута и расслаблена, возбуждаются мышечные рецепторы; Б — мышца сокращена (напряжена) и укорочена, возбуждаются сухожильные рецепторы; 1 — мышечные рецепторы (мышечные веретена); 2 — сухожилия мышцы и их рецепторы (рецепторы Гольджи); 3 — полусегмент спинного мозга
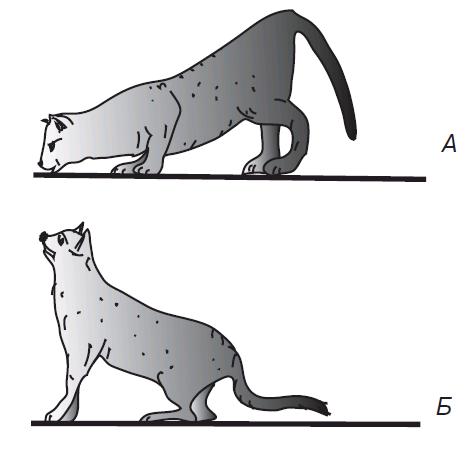
Рис. 4.15. Позные шейные рефлексы у кошки с удаленным вестибулярным аппаратом: А — при пассивном опускании; Б — при пассивном поднятии головы
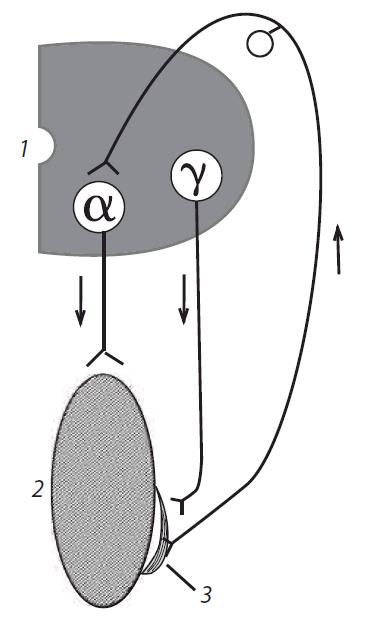
Рис. 4.16. Механизм регуляции тонуса мышц на уровне спинного мозга и развития гипертонуса мышц нижних конечностей в случае повреждения спинного мозга: 1 — полусегмент спинного мозга; 2 — скелетная мышца; 3 — мышечное веретено; α — мотонейрон, иннервирующий миоциты скелетной мышцы; γ — мотонейрон, иннервирующий миоциты мышечного рецептора
Двигательные системы ствола мозга
Характеристика мезэнцефального организма. Ствол мозга включает продолговатый мозг, мост и средний мозг (рис. 4.17), где локализуются ядра черепных нервов продолговатого мозга (IX–XII пары), моста (V–VIII пары) и среднего мозга (III, IV пары), а также другие ядра, управляющие мускулатурой туловища и конечностей. Перечислим черепные нервы: добавочный нерв (XI, n. accessories), языкоглоточный (IX, n. glossopharyngeus), подъязычный (XII, n. hypoglossus), преддверно-улитковый (VIII, n. vestibulocochlearis), блуждающий (X, n. vagus), лицевой (VII, n. facialis), отводящий (VI, n. abducens), тройничный (V, n. trigeminus), глазодвигательный (III, n. oculomotorius), блоковой (IV, n. trochlearis).
У мезэнцефального организма тонус мышц нормальный, он может сохранять позу и восстанавливать ее при нарушении, но произвольные движения отсутствуют. Центры ствола мозга обеспечивают регуляцию тонуса мышц и функции внутренних органов.
Все центры ствола мозга целесообразно объединить в три группы:
- жизненно важные вегетативные центры регуляции систем: дыхания, сердечно-сосудистой, пищеварения. Поражение продолговатого мозга ведет к остановке дыхания и смерти;
- центры защитных рефлексов, в осуществлении которых участвуют по несколько черепных нервов, — это рефлексы чихания, кашля, мигания, слезоотделения, рвоты;
- ядра ствола мозга, управляющие мускулатурой конечностей и туловища, которые включают несколько ядер:
– ядра шва (продолговатый мозг). Аксоны его нейронов поступают в спинной мозг, многие из них содержат серотонин. На интернейронах заднего рога этот путь образует тормозные синапсы (угнетение передачи сигналов), а на мотонейронах — возбуждающие. В целом моноаминонергические пути модулируют реактивность нейронных цепей спинного мозга, а не конкретные движения;
– вестибулярные ядра (мост) оказывают возбуждающее влияние на α- и γ-мотонейроны спинного мозга мышц-разгибателей конечностей, туловища, шеи и тормозное влияние на мотонейроны сгибателей этих частей тела. Подобный эффект вызывают и ретикулярные ядра моста (медиальный ретикулоспинальный путь);
– красные ядра (средний мозг, руброспинальный путь), напротив, оказывают тормозное влияние на мышцы-разгибатели и возбуждающее — на мышцы-сгибатели. Такое же влияние оказывает и рети-
кулярное гигантоклеточное ядро продолговатого мозга (латеральный ретикулоспинальный путь).
В случае нарушения соотношения возбуждающих и тормозных влияний на мотонейроны спинного мозга (нарушение связи продолговатого и среднего мозга) развивается децеребрационная ригидность (рис. 4.18).
У мезэнцефального организма при внезапных раздражениях возникает сторожевая реакция (ориентировочный рефлекс).
При действии света наблюдаются поворот головы и глаз в сторону света, фиксация взора, слежение за движущимся объектом (осуществляются с помощью верхних холмиков четверохолмия — первичного зрительного центра).
При действии звукового раздражителя наблюдаются прислушивание, поворот ушных раковин у животных (реализуются с помощью нижних холмиков четверохолмия — первичного слухового центра).
В обоих случаях наблюдается готовность к действию — повышается тонус мышц-сгибателей конечностей. Ориентировочный рефлекс — это запрограммированная реакция, как и рефлексы шагания, сосания, глотания.
Двигательные системы ствола мозга обеспечивают с помощью тонических рефлексов сохранение естественной позы и восстановление позы при ее нарушении. Рефлексы возникают с вестибулярных, проприоцептивных, тактильных и зрительных рецепторов и осуществляются двигательными центрами ствола мозга, включая ядра ретикулярной формации. Афферентные (восходящие) пути см. в табл. 4.1.
Эфферентные влияния из двигательных центров ствола мозга распространяются на мускулатуру конечностей и туловища по четырем нисходящим путям (табл. 4.1). Механизм возникновения децеребрационной ригидности и сохранения нормального тонуса мышц на уровне среднего мозга (мезэнцефальный организм) показан на рис. 4.19.
Классификация тонических (позных) рефлексов организма по Р. Магнусу с изменениями. Все тонические рефлексы (рефлексы позы) осуществляются с помощью одних и тех же двигательных ядер — красных, вестибулярных и ретикулярных ядер ствола мозга, ядер спинного мозга — и отличаются друг от друга только рефлексогенными зонами. Позные рефлексы включают три группы: статические, статокинетические и кинетические.Они выражаются в перераспределении мышечного тонуса.
Статические рефлексы (от греч. statos — стоящий) — это тонические рефлексы, обеспечивающие сохранение естественной позы организма в покое. Человек стоит или сидит, в том числе и при поворотах или наклонах головы и движениях рук, а сидя — и ног, вызывающих смещение центра тяжести. К ним относятся сегментарные и надсегментарные (шейные) тонические рефлексы спинного мозга и рефлексы ствола мозга, возникающие с проприо- и вести- булорецепторов. При вертикальной позе человека (голова вверх, шея наклонена несколько вперед) шейные и вестибулярные рефлексы усиливают друг друга, в результате чего происходит повышение тонуса разгибателей нижних конечностей и сгибателей верхних конечностей.
Статические тонические рефлексы открыл Бронжест в опыте на спинальной лягушке. Онперерезал задние корешки спинного мозга и подвесил лягушку на крючок. Нижняя конечность на стороне перерезки корешков была выпрямлена, а другая конечность слегка согнута, что свидетельствует о наличии рефлекторного тонуса мышц (см. раздел 4.10).
Статокинетические рефлексы —этотоническиерефлек- сы, обеспечивающие восстановление нарушенной позы. Они возникают с кожных, проприо- (шея) и вестибулорецепторов, при нарушении позы (организм находится в горизонтальном положении). К ним относятся выпрямительные (установочные) рефлексы — выпрямление головы и туловища.
Выпрямление головы запускается с вестибулярных и кожныхрецепторов, выпрямлениетуловища—с проприо- рецепторов шеи (они раздражаются при движении головы) и кожных рецепторов — по две рефлексогенных зоны. Именно поэтому при выключении только рецепторов кожи или вестибулярного аппарата у мезэнцефального организма рефлекс выпрямления головы не нарушается. При выключении же обеих названных рефлексогенных зон этот рефлекс не возникает. Рефлекс выпрямления туловища также исключается, если выключить проприорецепторы шеи (накладывание гипса на шею) и рецепторы кожи. Если одна из названных зон сохраняется, выпрямление туловища не нарушается. Вестибулярный аппарат в эксперименте выключается путем его разрушения или анестезией. Это надсегментарные рефлексы ствола мозга и спинного мозга. Особо важную роль играют красные ядра — при их разрушении нарушенная поза животного не восстанавливается.
Р. Магнус назвал эти рефлексы статическими. Однако они имеют две составляющих — статическую (нет перемещения тела в пространстве или вращения тела вокруг вертикальной оси на месте) и кинетическую (от греч. kinetikos — относящийся к движению) — животное поднимается на ноги и принимает естественную позу. У человека и обезьян выпрямительные рефлексы осуществляются при обязательном участии коры большого мозга, при ее выключении выпрямительные рефлексы не возникают.
Кинетические рефлексы (от греч. kinema — движение) — это тонические рефлексы, обеспечивающие сохранение естественной позы в движении — при ходьбе и беге, вращательном движении на месте, при изменении скорости движения в транспорте и в лифте. Поэтому называть их статокинетическими нет оснований — никакой статики, только движение. Эти рефлексы возникают с проприо- и вестибулорецепторов. Сокращения мышц направлены на преодоление сил гравитации, изменяющихся по величине вследствие ускорения или замедления движения. Важную роль играет система зрения. В процессе вращательных движений наблюдаются также повороты головы и нистагм глаз (пилообразные движения глаз).
Глазной нистагм способствует сохранению зрительной ориентации. Нистагм имеет две фазы. При вращении тела сначала происходит медленное отклонение глазных яблок в сторону, противоположную вращению тела. Затем глаза быстро движутся обратно — по ходу вращения «догоняют организм». Медленное отклонение вызывается с рецепторов полукружных каналов, быстрый компонент связан с влиянием корковых центров (при наркозе он исчезает). Во врачебной практике направление нистагма обозначают по быстрому компоненту. Исследование нистагма (степени его выраженности, длительности) используется для оценки функционального состояния вестибулярной системы и дифференциальной диагностики поражения ЦНС.
Лифтные рефлексы. При остановке движущегося вниз лифта и в начале его быстрого подъема в нижних конечностях повышается тонус мышц-разгибателей, что обеспечивает преодоление сил инерции хотя наблюдается некоторое сгибание конечностей под влиянием массы тела. В начале опускания лифта и при остановке движущегося вверх лифта наблюдаются противоположные явления. Существует и другая точка зрения, учитывающая только внешние признаки — пассивное сгибание или разгибание конечностей вследствие действия сил гравитации.
Таким образом, ствол мозга обеспечивает сохранение естественной позы, восстановление позы при ее нарушении, нормальный тонус мышц.
Совокупность проприоцептивной и вестибулярной сенсорных систем и ядер ствола мозга и спинного мозга образует систему положения тела.
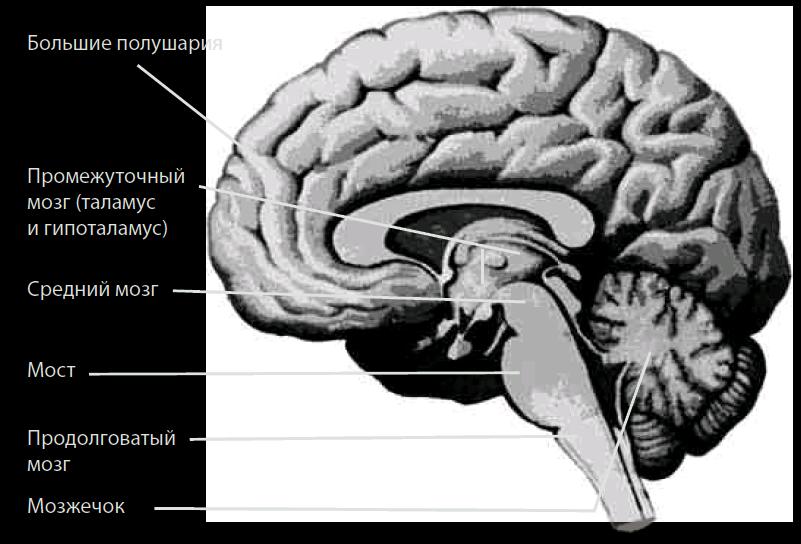
Рис. 4.17. Основные отделы головного мозга
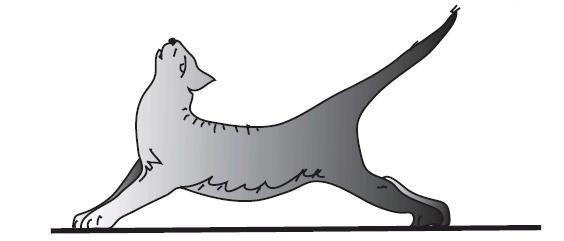
Рис. 4.18. Децеребрационная ригидность у кошки при перерезке ствола мозга между мостом и средним мозгом
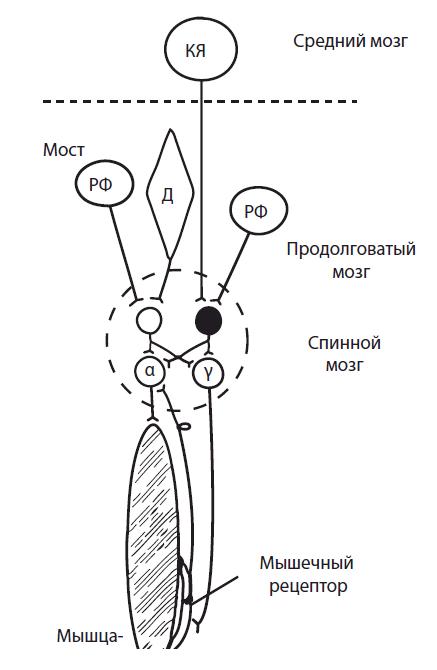
Рис. 4.19. Механизмы регуляции тонуса мышц и возникновения децеребрационной ригидности при перерезке ствола мозга между средним мозгом и мостом (пунктирная линия): РФ — ретикулярная формация; Д — ядро Дейтерса; α, γ — мотонейроны; вставочные нейроны спинного мозга: светлый — возбуждающий; темный — тормозной; КЯ — красное ядро
Интегративные системы ствола мозга
Интегративные системы ствола мозга включают ретикулярную формацию (РФ), голубое пятно, ядра шва, центральное серое вещество и черную субстанцию. Эти структуры имеют обширные нисходящие и восходящие связи со всеми отделами ЦНС.
Ретикулярная формация представляет собой скопления нейронов, различных по функции и размерам, связанных множеством нервных волокон, проходящих в разных направлениях и образующих сеть на всем протяжении ствола мозга, что и определяет ее название. Нейроны либо расположены диффузно, либо образуют ядра. Каждый нейрон РФ устанавливает связь с множеством других нейронов ЦНС (до 25 000). Нейроны ретикулярной формации имеют длинные маловетвящиеся дендриты и хорошо ветвящиеся аксоны, которые часто образуют Т-образное ветвление одна из ветвей аксона имеет нисходящее, а вторая — восходящее направление.
Свойства нейронов РФ:
- обладают спонтанной активностью, частота их разрядов в покое равна 5–10 имп./с;
- полисенсорность — они принимают коллатерали от нескольких сенсорных путей, идущих от разных рецепторов;
- им свойственна высокая чувствительность к некоторым веществам крови (например, адреналину, СО2) и лекарствам (барбитуратам, аминазину и др.);
- ониобладают высокой лабильностью —до 400имп./с;
- нейроны РФ более возбудимы по сравнению с другими нейронами.
Нейроны и ядра РФ входят в состав центров, регулирующих функции внутренних органов (кровообращения, дыхания, пищеварения), регулируют тонус скелетной мускулатуры, активность коры большого мозга. Обширны связи ретикулярной формации с другими отделами ЦНС и рефлексогенными зонами — РФ получает импульсацию от различных сенсорных рецепторов организма и отделов ЦНС и, в свою очередь, посылает импульсы во все отделы мозга. При этом выделяют восходящие и нисходящие влияния ретикулярной формации.
Нисходящие влияния РФ на моторные спинальные центры показаны на рис. 4.12 (мышцы-разгибатели). На сгибатели влияния противоположные. С помощью нисходящих влияний на спинной мозг РФ принимает участие в регуляции тонуса мышц, в формировании всех двигательных актов.
Восходящие влияния РФ на большой мозг могут быть как активирующими (они преобладают), так и тормозными. Импульсы от ретикулярных нейронов поступают к неспецифическим ядрам таламуса и после переключения в них проецируются в различные области коры, в задний гипоталамус, стриатум.
Вегетативные функции РФ заключаются в поддержании тонуса вегетативных центров, интеграции симпатических и парасимпатических влияний для реализации потребностей организма, в передаче модулирующих влияний от гипоталамуса и мозжечка к органам. РФ является главным структурным элементом жизненно важных центров продолговатого мозга — сердечно-сосудистого, дыхательного, жевательного, глотательного.
Черная субстанция участвует в регуляции мышечного тонуса, координации жевания и глотания, мелких движений конечностей (например, при печатании на машинке, письме), в осуществлении эмоций. В случае повреждения черной субстанции наблюдается бедность движений. Ее нейроны — дофаминергические, поэтому черную субстанцию выделяют как дофаминергическую систему.
Голубое пятно (ГП) расположено в среднем мозге, его нейроны являются адренергическими, как и вентролатерального ядра ретикулярной формации продолговатого мозга. В совокупности они составляют адренергическую систему ствола мозга. Нейроны ГП имеют фоновую импульсную активность (10–20 Гц). На их мембранах имеются рецепторы к адреналину, норадреналину, серотонину, ГАМК, глицину, опиоидам, активация которых вызывает преимущественно торможение, а также рецепторы к ацетилхолину, пептиду Р, глутамату, активация которых вызывает преимущественно возбуждение.
В свою очередь, нейроны ГП оказывают тормозное влияние на другие нейроны посредством β-адренорецепторов, реже — возбуждающее (например, в ядрах VII черепного нерва), посредством α-адренорецепторов. Импульсная активность нейронов голубого пятна повышена в фазе быстрого сна, что выключает мышечный тонус и фазические движения мускулатуры туловища и конечностей.
Иннервируя ядра гипоталамуса, нейроны ГП участвуют в развитии общего адаптационного синдрома при действии на организм стрессоров. Норадренергическая система контролирует болевую и неболевую импульсацию сенсорных и двигательных систем, угнетая фоновую активность нейронов их ядер. Оно участвует в регуляции деятельности висцеральных систем, эмоциональных состояний (тоска, страх), механизмов памяти и внимания, индуцирует фазу быстрого сна.
Серотонинергическая система ЦНС включает центральное серое вещество и ретикулярные магноцеллюляр- ные ядра, ядра шва, имеющие афферентные и эфферентные связи со всеми отделами ЦНС. Ее нейроны наиболее активны во время бодрствования, менее активны во время медленного сна, их возбуждение прекращается в фазе быстрого сна. Эта система тормозит нейроны сенсорных, особенно болевых, путей спинного мозга, ствола и таламуса. Серотонинергические нейроны тормозят агрессивное поведение, потребление воды и пищи.
Двигательные функции мозжечка
Структурно-функциональная характеристика мозжечка. Главная функциия мозжечка — координация автоматизированных движений. Мозжечок содержит больше половины всех нейронов ЦНС, но его масса составляет всего лишь 10% массы головного мозга. Выделяют три основных структуры мозжечка: древний мозжечок, старый мозжечок и новый мозжечок.
Афферентные входы и эфферентные выходы мозжечка осуществляются посредством ножек: нижних — с продолговатым мозгом (афферентная импульсация в мозжечок поступает от вестибуло- и проприорецепторов), средних — с мостом и корой большого мозга, верхних — со средним мозгом (рис. 4.20).
Роль мозжечка в организации движений. Мозжечок участвует в регуляции тонуса мышц — он оказывает тормозное влияние посредством красных и вестибулярных ядер, ретикулярной формации ствола мозга. Поддержание равновесия (позы) осуществляется с помощью перераспределения мышечного тонуса. Этому способствует импуль- сация от вестибулярных рецепторов, проприорецепторов, кожных, зрительных и слуховых рецепторов. Наиболее характерным симптомом поражения древнего мозжечка является нарушение равновесия.
Координация выполняемых движений реализуется старым и новым мозжечком. Сюда поступает информация от проприорецепторов и от моторной коры большого мозга (программа движения). Координация достигается с помощью анализа этой информации и реализуется через свои промежуточные ядра, имеющие выходы на красное ядро и моторную кору. При этом регулируются амплитуда, направление и темп движения. Наиболее характерным симптомом нарушения функции промежуточной зоны мозжечка является нарушение координации движения (атаксия). Еще одна функция мозжечка — коррекция быстрых движений, которая из-за большой скорости их развития не может быть осуществлена в процессе движения с помощью импульса- ций от проприорецепторов. Мозжечок получает копию эфферентной программы движения от моторной коры большого мозга до начала движения. Кора мозжечка обучена в фило- и онтогенезе исправлять возможные ошибки, с учетом которых изменяется эфферентная программа движения (т.е. мозжечок выполняет роль, аналогичную акцептору результата действия в функциональной системе). К таким движениям относятся многие быстрые спортивные движения, игра на музыкальных инструментах и др.
Программирование целенаправленных движений осуществляется корой нового мозжечка, которая получает импульсацию преимущественно из ассоциативных зон коры большого мозга через ядра моста. Эта информация характеризует замысел движения. Инициация движения обеспечивается лимбической системой и ассоциативной корой. В коре нового мозжечка (а также в базальных ядрах, в моторной коре) она перерабатывается в программу движения и через таламус поступает в премоторную кору. Там осуществляется дальнейшая обработка информации, и через пирамидную и экстрапирамидную системы она реализуется в виде сложного целенаправленного движения.
Вегетативные функции мозжечка заключаются в том, что он, как и гипоталамус, участвует в согласовании вегетативного обеспечения соматической деятельности организма (модулирующее влияние на деятельность внутренних органов) посредством РФ, вегетативной нервной системы и эндокринной системы. После удаления мозжечка в эксперименте гомеостазис становится неустойчивым.
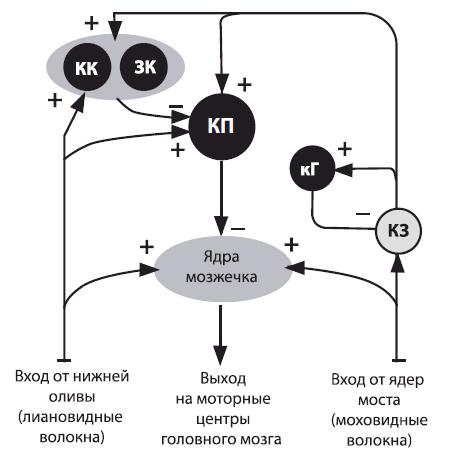
Рис. 4.20. Схема основных межнейронных связей мозжечка (по В.Н. Яковлеву): КП — клетки Пуркинье; КЗ — клетки зерна; КК — корзинчатые клетки; ЗК — звездчатые клетки; кГ — клетки Гольджи; «+», «–» — возбуждающие и тормозные влияния
Функции промежуточного мозга
Промежуточный мозг (diencephalon) расположен между средним и конечным мозгом. Он включает таламическую область (таламус, метаталамус и эпиталамус — эпифиз) и гипоталамус.
Таламус (thalamus — зрительный бугор) и метаталамус (коленчатые тела). Таламус представляет собой парный ядерный комплекс, включающий до 60 ядер, которые подразделяют на несколько групп: релейные (переключательные — сенсорные и несонсорные), специфические и неспецифические и ассоциативные. Изобилие терминов затрудняет усвоение материала. В частности термины «специфические» и «неспецифические» — некорректные. Каждое ядро или центр выполняет свою специфическую функцию. Поэтому термины «специфические» и «неспецифические» ядра целесообразно исключить. По функции основные ядра таламуса следует объединить в две группы переключательные — сенсорные и двигательные (релейные, реле-переключатель) — и ассоциативные (объединяющие, интегрирующие). Причем, переключательные ядра не только посылают информацию к коре большого мозга, но и получают обратные входы от соответствующих зон коры большого мозга, обеспечивающих взаимодействие между таламусом и корой большого мозга.
Переключательные ядра таламуса
Переключательные сенсорные ядра (вентральные задние ядра) направляют потоки афферентной импульсации в сенсорные зоны коры большого мозга. В них переключаются тактильная, проприоцептивная, вкусовая, висцеральная, частично температурная и болевая импульсации. В этих ядрах имеется топографическая проекция периферии; при этом функционально более тонко организованные ,части тела (на- пример,язык,кисть)имеютбольшуюзонупредставительства. В таламусе начинают формироваться ощущения — электростимуляция вентральных задних ядер вызывает парестезии (ложные ощущения) в разных частях тела. Импульсация от вентральных задних ядер проецируется в соматосенсорную кору постцентральной извилины, где формируются соответствующие ощущения. В латеральном коленчатом теле переключается зрительная импульсация и направляется в затылочную кору. Медиальное коленчатое тело переключает слуховую импульсацию, поступающую в височную кору.
Переключательные двигательные ядра таламуса (вентральные) направляют программы движений, сформированные в базальных ядрах и в мозжечке, в двигательную кору большого мозга (поля 4 и 6).
Ассоциативные ядра таламуса — это, во-первых, ядра РФ (ретикулярные ядра). Они имеют многочисленные входы от других ядер головного мозга, сенсорных систем и коры большого мозга. В свою очередь, они посылают импульсы ко всем структурам ЦНС. Благодаря этим связям ретикулярные ядра таламуса выступают в роли интегрирующего посредника между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ядрами — с другой, объединяя их в единый функциональный комплекс. На кору мозга они оказывают преимущественно модулирующее (изменяющее состояние) влияние. Поэтому нарушение его функции не вызывает грубых расстройств. Во-вторых, ассоциативными являются группа ядер таламуса (подушка, медиодорсальное ядро, латеральные ядра — ассоциативные ядра таламуса), которые получают импульсацию от других ядер самого таламуса, от зрительных и слуховых путей и других ядер головного мозга и направляют ее в основном в ассоциативные зоны коры большого мозга. В свою очередь, кора большого мозга посылает импульсы к ассоциативным ядрам, регулируя их интегративную функцию.
Гипоталамус включает преоптическую область, сосцевидные (мамиллярные) тела, область перекреста зрительных нервов, серый бугор и воронку. В гипоталамусе выделяют несколько десятков парных ядер, которые объединяют в несколько групп.
Связи гипоталамуса. Его ядра образуют многочисленные связи друг с другом, а также с выше- и нижележащими структурами ЦНС. Главные входы в гипоталамус: от лимбической системы, коры большого мозга, базальных ядер и ретикулярной формации ствола мозга. Основные выходы гипоталамуса: в ствол мозга (ретикулярную формацию, моторные и вегетативные центры), в вегетативные центры спинного мозга, в лимбическую систему, от супраоптического и паравентрикулярного ядер — к нейрогипофизу, от вентромедиального и аркуатного ядер — к аденогипофизу, а также к стриатуму и лобной коре.
Функции гипоталамуса. Он является многофункциональной системой, обладающей широкими регулирующими и интегрирующими влияниями в регуляции деятельности внутренних органов, обмена веществ и энергии. При этом обычно отдельно взятое ядро выполняет несколько функций, а отдельно взятая функция локализуется в не-
скольких ядрах. Свои влияния на функции внутренних органов гипоталамус осуществляет посредством гормонов эндокринных желез, симпатической (задние ядра) и парасимпатической (передние ядра) систем.
Функции эпифиза описаны в разделе 6.5.
Функции базальных ядер и лимбической системы
Основные структуры базальных ядер (стриопаллидар- ная система): полосатое тело (corpus striatum), называемое также striatum (хвостатое ядро и скорлупа), бледный шар и субталамическое ядро.
Двигательные функции базальных ядер. В целом базальные ядра, имея двусторонние связи с корой большого мозга, таламусом, ядрами ствола мозга, участвуют в создании программ целенаправленных движений с учетом доминирующей мотивации. При этом нейроны стриатума оказывают тормозное влияние (медиатор — ГАМК) на нейроны черного вещества. В свою очередь, нейроны черного вещества (медиатор — дофамин) оказывают модулирующее влияние (тормозное и возбуждающее) на фоновую активность нейронов стриатума. При нарушении дофаминергических влияний на базальные ядра наблюдаются двигательные расстройства типа паркинсонизма, при которых резко падает концентрация дофамина в обоих ядрах стриатума. Наиболее важные функции базальных ядер выполняют стриатум и бледный шар.
Функции стриатума. Участвует в осуществлении поворота головы и туловища и ходьбы по кругу, которые входят в структуру ориентировочного поведения. Поражение хвостатого ядра при заболеваниях и при разрушении в эксперименте ведет к насильственным, избыточным движениям (гиперкинезы: хорея и атетоз).
Функции бледного шара. Оказывает модулирующее влияние на двигательную кору, мозжечок, РФ, красное ядро. При стимуляции бледного шара у животных преобладают элементарные двигательные реакции в виде сокращения мышц конечностей, шеи и лица, активация пищевого поведения. Разрушение бледного шара сопровождается снижением двигательной активности — возникает адинамия (бледность двигательных реакций), а также развитие сонливости, «эмоциональной тупости», что затрудняет реализацию имеющихся условных рефлексов и ухудшает выработку новых (ухудшает кратковременную память).
Структурно-функциональная организация лимбической системы (от лат. limbus — край) — это функциональное объединение различных структур, среднего, промежуточного и конечного мозга, обеспечивающее мотивацию поведенческих реакций и их вегетативное обеспечение. Мотивации формируют также эмоциональную окраску поведения.
Структуры. Корковые: обонятельная извилина, гиппокамп (аммонов рог), зубчатая, поясная и парагиппокампальная извилины; подкорковые: миндалевидное тело, область перегородки, ограда, гипоталамус, переднее таламическое ядро.
Входы в лимбическую систему осуществляются от различных областей головного мозга, от обонятельных рецепторов по волокнам обонятельного нерва (I, n. olfactorii). Главным источником возбуждения лимбической системы является РФ ствола мозга. Выходы из лимбической системы осуществляются через гипоталамус к нижележащим вегетативным и соматическим центрам ствола мозга и спинного мозга (нисходящие влияния). Восходящая импульсация в новую кору направляется преимущественно в ассоциативные области. Кольцевые нейронные связи объединяют различные ее структуры и делают возможным длительную циркуляцию (реверберацию) возбуждения, которая является механизмом его пролонгирования (рис. 4.21).
Функции лимбической системы. Получая информацию о внешней и внутренней средах организма, лимбическая система, после сравнения и обработки этой информации, запускает вегетативные и соматические (поведенческие) реакции, обеспечивающие адекватное приспособление организма к внешней среде и сохранение гомеостазиса. Частные функции лимбической системы:
- регуляция функций внутренних органов, осуществляется преимущественно через гипоталамус;
- формирование мотиваций, эмоций и поведенческих реакций (агрессивно-оборонительных, пищевых и сексуальных;
- играет важную роль в обучении.
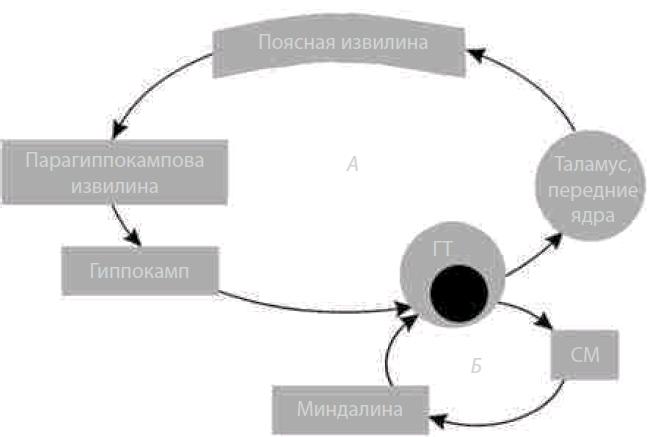
Рис. 4.21. Схема основных внутренних связей лимбической системы (по В.Н. Яковлеву): А — круг Пейпеца; Б — круг через миндалину; ГТ/МТ — мамиллярные тела гипоталамуса; СМ — средний мозг (лимбическая область)
Нейронная организация новой коры. Функциональные зоны
Новая кора (неокортекс) — это слой серого вещества, общая площадь которого за счет складок достигает 2000 см2, она покрывает большие полушария и составляет около 70% всей площади коры.
Общая характеристика коры. В новой коре имеются поли- и монофункциональные поля. Свойство полифункциональности позволяет данной корковой структуре включаться в обеспечение различных форм деятельности, реализуя при этом основную, генетически присущую ей функцию (Андрианов О.С., 1976). Степень полифункциональности различных корковых структур неодинакова например, поля ассоциативной коры полисенсорны, первичные сенсорные зоны моносенсорны, во вторичных сенсорных зонах локализуются преимущественно бисенсорные нейроны. В основе мультифункциональности лежит многоканальность поступления в кору мозга афферентного возбуждения. Детальное деление коры на различные поля проведено на основе цитоархитектонических признаков (формы и расположения нейронов) К. Бродманом (1909), который выделил 52 поля, многие из них характеризуются функциональными и нейрохимическими особенностями. В направлении с поверхности вглубь неокортекс имеет шесть горизонтальных слоев.
В слоях I и IV новой коры происходят восприятие и обработка поступающих сигналов. Нейроны II и III слоев осуществляют кортикокортикальные ассоциативные связи. Нейроны V и VI слоев формируют нисходящие пути.
Функциональные нейронные колонки новой коры большого мозга. В коре мозга имеются функциональные объединения нейронов, расположенные в цилиндрике диаметром 0,5–1,0 мм, включающем все слои коры и содержащем несколько сотен нейронов (нейронные колонки). Об этом, в частности, свидетельствуют электрофизиологические исследования с погружением микроэлектродов перпендикулярно поверхности соматосенсорной коры. При этом все встречаемые на пути нейроны отвечают на раздражитель только одного вида (например, свет). При погружении электрода под углом на его пути попадались нейроны разной сенсорности. Колонки обнаружены в моторной коре и различных зонах сенсорной коры. Нейроны колонки могут осуществлять саморегуляцию по типу возвратного торможения. Соседние нейронные колонки могут частично перекрываться, а также взаимодействовать друг с другом по механизму латерального торможения.
Кортикализация функций — это возрастание в филогенезе роли коры большого мозга в регуляции функций организма и подчинение нижележащих отделов ЦНС, в обеспечении психической деятельности организма. Например, регуляция локомоторных двигательных функций (прыжки, ходьба, бег) и выпрямительных рефлексов у низших позвоночных полностью обеспечивается стволом мозга, удаление больших полушарий практически их не изменяет. У кошек после перерезки ствола между средним и промежуточным мозгом локомоция сохраняется лишь частично. Выключение коры большого мозга в эксперименте у обезьян и в патологических случаях у человека ведет к потере не только произвольных движений, но и выпрямительных рефлексов.
Функциональные зоны новой коры. Функциональными отделами коры большого мозга являются сенсорные, ассоциативные и двигательные зоны (рис. 4.22).
Сенсорные зоны коры (проекционная кора, корковые отделы сенсорных систем). Характеристика сенсорной коры. В эти зоны поступает информация от сенсорных рецепторов. Они расположены в теменной, височной и затылочной долях. Афферентные пути в сенсорную кору поступают преимущественно от специфических сенсорных ядер таламуса. Участки сенсорной коры, раздражение или разрушение которых вызывает четкие и постоянные изменения определенной чувствительности организма, называют первичными сенсорными зонами. Они состоят преимущественно из моносенсорных нейронов и формируют ощущения одного вида (качества). В первичных сенсорных зонах обычно имеется четкое пространственное (топографическое) представительство частей тела, от которых поступает импульсация (рис. 4.23).
Вокруг первичных сенсорных зон локализуются вторичные сенсорные зоны,нейроны которых преимущественно бисенсорные — они реагируют обычно на два раздражителя, например на свет и звук. Во вторичные сенсорные зоны поступает также информация от первичных, что способствует формированию образа объекта или явления.
Главные сенсорные зоны
- Соматосенсорная область (SI) — это постцентральная извилина теменной доли и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий (поля 1–3). При этом поле 3 представляет собой первичное сенсорное поле, а поля 1–2 — вторичные сенсорные поля. В области SI имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной и проприоцептивной чувствительности. Проекции наиболее чувствительных участков (язык, губы, гортань, пальцы рук) имеют относительно большие зоны по сравнению с другими частями тела. Предполагают, что в зоне тактильной чувствительности языка расположена проекция и вкусовой чувствительности. Имеются также меньших размеров вторичная соматосенсорная область (SII), расположенная в стенке боковой борозды, на границе ее пересечения с центральной бороздой. Лобная и теменная доли разделены центральной бороздой. Зона SII, по-видимому, участвует в сенсорной и моторной координации двух сторон тела, локализация поверхности тела в ней менее четкая, чем в области SI.
- Височная область — здесь в верхней височной извилине расположена слуховая сенсорная кора. Первичной сенсорной зоной является поле 41, в глубине боковой борозды с четкой топической проекцией различных участков кортиева органа. Вторичная сенсорная зона расположена в верхней височной извилине (поле 42 ), где происходит более сложная обработка звуковой (частично и речевой) информации;
- Затылочнаяобласть (поле 17),где локализуетсяпервич- ная сенсорная зона зрительной системы с топическим представительством рецепторов сетчатки, особенно желтого пятна (основная часть поля 17), рядом располагается вторичная сенсорная зона (поля 18, 19). Обработанная в первичной и вторичной сенсорной коре информация передается для дальнейшей обработки в ассоциативную кору — третичную сенсорную зону.
Взаимодействие трех сенсорных зон обеспечивает с помощью мышления формирование комплексного восприятия и формирование соответствующего поведения.
Ассоциативные области коры. Следует заметить, что описание ассоциативных областей несколько различается у разных авторов.
Теменная ассоциативная кора (поля 5, 7, 40) получает основные афферентные входы от задней группы ассоциативных ядер таламуса, имеет эфферентные выходы на ядра таламуса и гипоталамуса, моторную кору и ядра экстрапирамидной системы. Основными функциями теменной ассоциативной коры являются гнозис и прак- сис. Под гнозисом понимают функцию различных видов узнавания: формы, величины, значения предметов, узнавание их на ощупь, понимание речи, познание процессов, закономерностей и др. Под праксисом понимают целенаправленное автоматизированное действие (например, печатание на клавиатуре компьютера). Понимание сложных логико-грамматических конструкций обеспечивается нижнетеменными и теменно-затылочными областями коры большого мозга. Эти области обеспечивают ориентировку в пространстве.
Лобная (фронтальная) ассоциативная кора имеет основной афферентный вход от теменной ассоциативной коры и ассоциативного медиодорсального ядра таламуса. Главная функциия лобной ассоциативной коры — формирование программ целенаправленного поведения (особенно в новой для человека обстановке), под влиянием доминирующей мотивации, обеспечивающей направление поведения человека. Эта функция основана на тесных двусторонних связях лобной коры с лимбической системой. Причем лобная кора обеспечивает вероятностное прогнозирование, что выражается изменением поведения в ответ на изменения обстановки окружающей среды и доминирующей мотивации. В лобной коре осуществляется самоконтроль действий путем постоянного сравнения результата действия с исходными намерениями, что связано с созданием аппарата предвидения (акцептора) результата действия в функциональной системе по П.К. Анохину.
В результате проведения по медицинским показаниям префронтальной лоботомии, при которой пересекаются связи между лобной долей и таламусом, наблюдается развитие «эмоциональной тупости», отсутствие мотивации, твердых намерений и планов,основанных на прогнозировании. План поведения, формируемый ассоциативной корой, реализуется с помощью двигательной коры.
Некоторые авторы выделяют теменно-затылочно-височную ассоциативную область, функции которой в основном совпадают с вышеописанными, а также — лимбическую область, участвующую в формировании мотиваций, эмоций и поведения.
Двигательные зоны коры — это первичная двигательная (моторная) кора прецентральной извилины (поле 4) и вторичная двигательная кора (поле 6), которая включает прилегающую к ней рострально-премоторную область и дополнительную двигательную область на медиальной стороне полушария. Пирамидные нейроны V слоя этих областей (формирующие пирамидные пути) составляют двигательные колонки и иннервируют α- и γ-мотонейроны двигательных центров как непосредственно (моносинап- тически), так и через интернейроны двигательных центров (полисинаптически). Пирамидные нейроны двигательной колонки получают импульсы от проприо- и тактильных рецепторов, формируют двигательные команды и посылают их через нейроны верхних слоев к стволовым и спинальным центрам, последние управляют мышцами одного сустава или одной мышцы, но мышцы туловища могут иннервироваться от нескольких колонок. Соседние колонки в функциональном плане перекрываются. В двигательные области информация поступает по восходящим афферентам через таламус, непосредственно от таламуса, и интегрированная информация от корковых нейронов различных сенсорных систем, ассоциативной коры и лимбической системы.
Первичная двигательная кора. Ее пирамидные клетки, иннервируют мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела (рис. 4.24).
Нейроны коры управляют функцией мышц противоположной стороны тела. При поражениях этой корковой зоны нарушается способность к тонким координированным движениям конечностей, исключаются тонкие движения пальцев руки на противоположной стороне, движение языка и нижней части лица.
Вторичная двигательная кора имеет главенствующее значение по отношению к первичной двигательной коре и обеспечивает осуществление более сложных координированных движений, играет важную роль в планировании движения. Поражения вторичной двигательной зоны ведут к резкому ограничению движений у человека. В премоторной коре локализуется моторный центр речи (центр Брока), управления движениями глаз. При раздражении вторичной коры возникают более сложные движения (конечностей, туловища, головы), чем при раздражении первичной двигательной коры, например одного сустава, движения пальцев рук. Двигательная кора осуществляет свои функции через пирамидную и экстрапирамидную системы.
Пирамидная система — совокупность нейронов пирамидного пути (от первичной двигательной коры), α- и γ-мотонейронов ствола мозга и спинного мозга, синап- тически связанных друг с другом непосредственно и с помощью вставочных нейронов. Пирамидный путь образуют аксоны пирамидных клеток V слоя коры большого мозга (клетки Беца, первичная двигательная кора). Пирамидная система играет особо важную роль в выполнении тонких движений пальцев рук и кистей.
Экстрапирамидная система (ЭПС) — совокупность двигательных ядер головного мозга, большинство из которых не имеют прямой связи с α- и γ-мотонейронами спинного мозга и ствола мозга или со вставочными нейронами спинного мозга. Ядра ЭПС — двигательная кора (кроме первичной), базальные ядра, черная субстанция, красные ядра, люисово тело (субталамическое ядро), ретикулярная формация, центры четверохолмия, голубое пятно, вестибулярные ядра и мозжечок. Функция — регуляция мышечного тонуса, сохранение естественной позы, восстановление нарушенной позы, участие в осуществлении произвольных движений.
Межполушарные функциональные связи осуществляются через мозолистое тело, а также комиссуры (спайки), они играют важную роль в деятельности организма, что показано в эксперименте на животных с рассечением мозолистого тела и в результате подобной операции у пациентов по медицинским показаниям (чтобы прекратить межполушарное распространение эпилептических судорог). При этом возникают различные нарушения — двигательные, речевые, узнавания с помощью зрительной и тактильной систем. Такие больные могут рассказать о соматосенсорных стимулах, нанесенных на правую сторону тела, но не на левую, т.к. информация, поступающая в правые сенсомоторные области коры (от левой половины тела), не достигает речевых центров (они расположены в левом полушарии), но полушария могут выполнять отдельные функции независимо друг от друга. У здорового же организма мозг работает как единое целое.
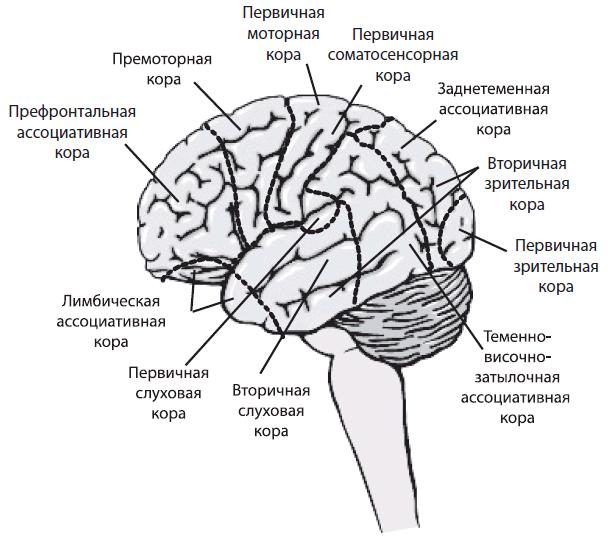
Рис. 4.22. Функциональные области коры большого мозга
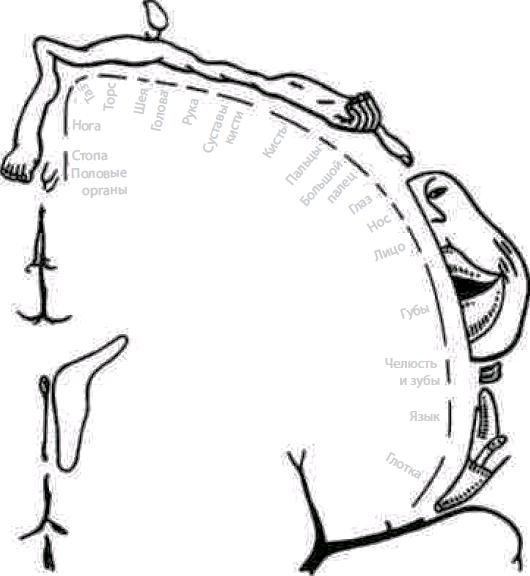
Рис. 4.23. Чувствительный гомункулус — проекция общей чувствительности в коре (постцентральная извилина) (по У. Пенфилду, Т. Расмуссену, 1950)
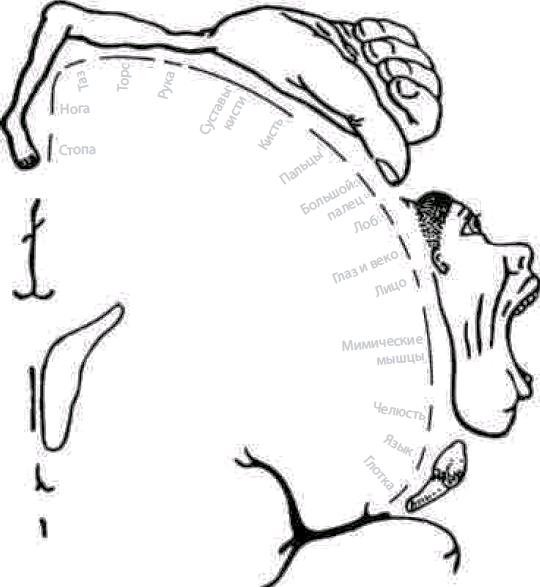
Рис. 4.24. Двигательный гомункулус — проекция двигательной системы в коре (прецентральная извилина) (по У. Пенфилду, Т. Расмуссену, 1950)
